
Thomas Duncan, PhD
CURRENT APPOINTMENTS
LANGUAGES
WEB RESOURCES
RESEARCH PROGRAMS AND AFFILIATIONS
RESEARCH INTERESTS
F-type ATP synthases; bioenergetics of pathogenic bacteria; enzymology; structural biology, membrane protein function.
EDUCATION INTERESTS
Bioenergetics; mitochondria; lipid and cholesterol metabolism
ASSOCIATIONS / MEMBERSHIPS
EDUCATION
RESEARCH ABSTRACT
My research focuses on the structure and function of the F-type ATP synthase, an energy-transducing enzyme complex that is critical to the energy metabolism of most living things. My current emphasis is on its critical roles in bacterial metabolism, virulence and pathogenesis. The F-type ATP synthase is assembled on the inner cell membrane of bacteria, the inner mitochondrial membrane of eukaryotes and on the thylakoid membrane of chloroplasts (see example for the ‘simple’ bacterial enzyme, Fig. 1). In the terminal step of oxidative- and photo-phosphorylation, it uses energy from a transmembrane, electrochemical gradient of protons (proton-motive force, or pmf) to drive the synthesis of most cellular ATP. However, the enzyme is intrinsically reversible and, if the driving force is lost for respiration (e.g., lack of O2) or photosynthesis (darkness), the enzyme can shift to hydrolyzing cellular ATP, driving proton (H+) transport across the membrane in the opposite direction and generating pmf. The enzyme is critical for the viability and/or virulence of diverse pathogenic bacteria, some of which rely on it for ATP synthesis while others can only use it as an ATPase-driven H+ pump. Despite the general structural/functional conservation between bacterial and eukaryotic forms of the enzyme, the potential to target the enzyme to attack pathogenic bacteria has been confirmed by the development of Bedaquiline, which was found to target mycobacterial FOF1 and is currently approved as part of a 3-drug, front-line regimen to treat multi-drug resistant Tuberculosis. Bedaquiline is the first in a new diarylquinoline class of antibiotics that is highly selective for mycobacterial FOF1.
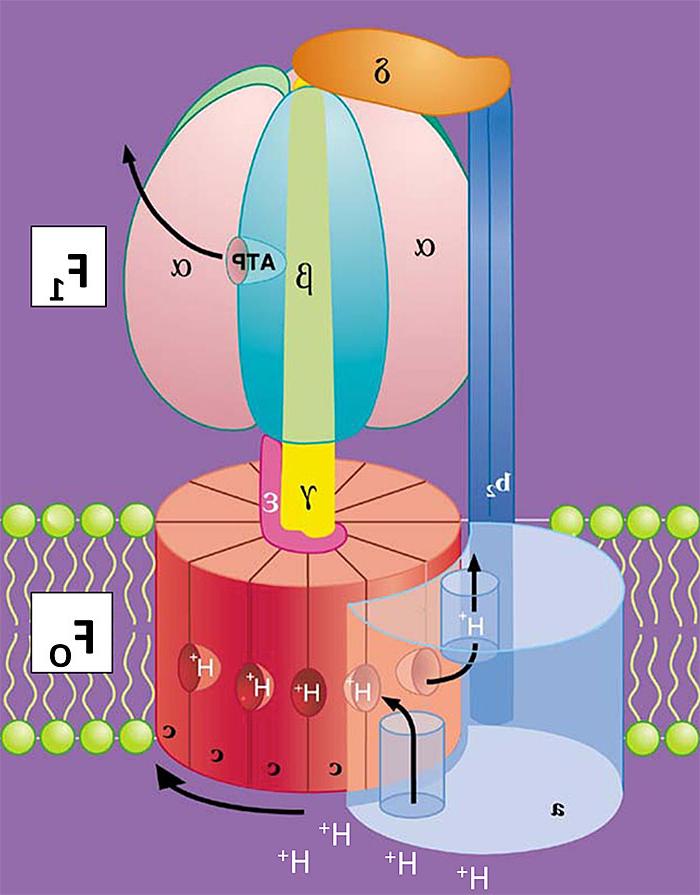
Fig. 1. Cartoon of E. coli FOF1 Architecture. Arrow below the c-ring shows direction the rotor complex (c10γε) turns during synthesis of ATP, as driven by H+ transport.
BACKGROUND
The ATP synthase consists of two sub-complexes with distinct but coupled functions. The basic architecture of the enzyme is shown in cartoon form in Fig. 1 for the minimal subunit composition found in most bacteria. The FO complex catalyzes H+ transport and has subunits that span the membrane bilayer. F1 is a peripheral complex that extends into the aqueous phase and contains three catalytic nucleotide-binding sites for synthesis and hydrolysis of ATP. FO and F1 are coupled through two stalk-like connections of subunits: a central rotor shaft and a peripheral stator. My earlier studies with Richard Cross provided key initial evidence that energy coupling by the ATP synthase involves
- Rotation of the central shaft within F1 to coordinate actions of the three cooperative, alternating catalytic nucleotide sites, and
- Rotation of subunits within FO during energy-driven proton transport.

Fig. 2. Minimal E. coli FOF1 Animation. Likely conformational transitions of one ß subunit and the εCTD relative to one small (~36°) rotary sub-step. One c-subunit is green to track rotation of the c-ring along with the rotor shaft of γ (yellow) and ε (NTD/CTD, pink/magenta). Only 1 other ß (cyan) is shown, and the intervening alpha subunits are not shown.
EXPERIMENTAL SYSTEMS & CURRENT PROJECTS
Our research has long used the bacterial ATP synthase from E. coli, which provides a simple yet powerful model for structure/function studies of this rotary motor enzyme. The E. coli system allows straightforward genetic screening and engineering of the ATP synthase, and also large-scale purification of FOF1 and isolated, soluble F1 for biochemical and structural studies. We continue to develop more detailed knowledge of E. coli FOF1 to understand the inhibitory mechanism of the εCTD and how to best target it to disrupt bacterial bioenergetics. For example, an ongoing collaboration with a single-molecule microscopist, Dr. Michael Börsch (Jena University, Germany) is using fluorescence resonance energy transfer (FRET) to study the dynamics of ε’s conformational changes in single EcFOF1-liposomes.
Much less is known about FOF1 from other pathogenic bacterial species, and we have begun to study the function and impact of FOF1 in 2 distinct pathogens:
- Pseudomonas aeruginosa. This gram-negative, opportunistic pathogen is a major contributor to persistent lung infections and mortality in cystic fibrosis patients. It is a substantial hazard for immune-compromised persons and is prevalent in hospital-acquired infections. Multi-drug-resistant strains continue to emerge and, in 2017, the World Health Organization listed carbapenem-resistant P. aeruginosa as one of three pathogens in critical need of new antibiotic development. P. aeruginosa metabolism is strongly dependent on respiration and the ATP synthase is essential for its viability. We are currently collaborating with the labs of Christopher Nomura at neighboring SUNY-ESF and Guirong Wang (Upstate Dept of Surgery) to study (i) how the εCTD of FOF1 impacts the enzyme’s function in P. aeruginosa in vitro and (ii) whether disruption of the εCTD impacts the virulence or pathogenesis of P. aeruginosa in cultured macrophages and in a mouse model of lung infection.
- Streptococcus pneumoniae. This gram-positive pathogen is the major cause of bacterial pneumonia but can also lead to severe invasive pneumococcal diseases such as sepsis and meningitis. This aerotolerant anaerobe lacks a complete respiratory chain, so it cannot use FOF1 for significant ATP synthesis. Nevertheless, FOF1 is essential for S. pneumoniae’s viability and acid tolerance, acting as an ATPase-driven H+ pump to generate pmf across the cell membrane and to maintain cellular pH homeostasis. A recent study of septicemia in mice, by the research group of Marco Oggioni (University of Leicester, UK), suggested a significant virulence role of pneumococcal FOF1: subclones of S. pneumoniae that emerged to establish septicemia most often contained a nascent missense mutation in a gene for one of the subunits of FOF1 (2014, Gerlini et al., PLoS Pathogens, 10 e1004026). One such virulent clone instead had a frame-shift mutation in the atpC gene for ε that ‘scrambled’ the sequence of the εCTD, and that pneumococcal clone showed increased survival in the spleen. They transferred that atpC mutation to a naïve, nonencapsulated pneumococcal strain, and we are working with it to determine how this disruption of the εCTD affects FOF1 function in pneumococcal membranes.
SELECTED REFERENCES
T.M. Duncan (2019). Turbine enzyme’s structure in the crosshairs to target tuberculosis. Proc. Natl. Acad. Sci. USA 116, 3956-58. http://doi.org/10.1073/pnas.1900798116
M. Sobti, R. Ishmukhametov, J.C. Bouwer, A. Ayer, C. Suarna, N.J. Smith, M. Christie, R. Stocker, T.M. Duncan, A.G. Stewart (2019). Cryo-EM reveals distinct conformations of E. coli ATP synthase on exposure to ATP. eLife 8, e43864. http://doi.org/10.7554/eLife.43864
H. Sielaff, T.M. Duncan, M. Börsch (2018). The regulatory subunit ε in Escherichia coli FOF1-ATP synthase. Biochim. Biophys. Acta Bioenerg. 1859, 775-788. http://doi.org/10.1016/j.bbabio.2018.06.013
N.B. Shah, T.M. Duncan (2015). Aerobic growth of Escherichia coli is reduced and ATP synthesis is selectively inhibited when five C-terminal residues are deleted from the ε subunit of ATP synthase, J. Biol. Chem. 290, 21032-41. http://doi.org/10.1074/jbc.M115.665059
M. Börsch, T.M. Duncan (2013). Spotlighting motors and controls of single FOF1-ATP synthase. Biochem. Soc. Trans. 41, 1219-26. http://doi.org/10.1042/BST20130101
N.B. Shah, M.L. Hutcheon, B.K. Haarer, T.M. Duncan (2013). F1-ATPase of Escherichia coli: The ε-Inhibited State Forms After ATP Hydrolysis, is Distinct from the ADP-Inhibited State, and Responds Dynamically to Catalytic-Site Ligands. J. Biol. Chem. 288, 9383-95. http://doi.org/10.1074/jbc.M113.451583
G. Cingolani, T.M. Duncan (2011). Structure of the ATP synthase catalytic complex (F1) from Escherichia coli in an autoinhibited conformation. Nat. Struc. Mol. Biol. 18, 701-7. http://doi.org/10.1038/nsmb.2058
T.M. Duncan (2004). The ATP Synthase: Parts and Properties of a Rotary Motor, in: The Enzymes, vol. XXIII: Energy Coupling and Molecular Motors, vol. 23, (D.D. Hackney, F. Tamanoi, Eds.) Elsevier Academic Press, New York, pp. 203-275.
M.L. Hutcheon, T.M. Duncan, H. Ngai, R.L. Cross (2001). Energy-driven subunit rotation at the interface between subunit a and the c oligomer in the FO sector of Escherichia coli ATP synthase. Proc. Natl. Acad. Sci.. USA 98, 8519-24. http://doi.org/10.1073/pnas.151236798
T.M. Duncan, V.V. Bulygin, Y. Zhou, M.L. Hutcheon, R.L. Cross (1995). Rotation of subunits during catalysis by Escherichia coli F1-ATPase. Proc. Natl. Acad. Sci. USA 92, 10964-68. http://doi.org/10.1073/pnas.92.24.10964